


Evidently, in our solar system life first evolved quite soon after the formation of a hospitable terrestrial environment. Suppose the typical time that it takes for life to evolve is denoted by some timescale tbio, then from the evidence presented by the solar system, which is about 4.6 x 109 yrs old, it is seems that
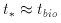
At first sight we might assume that the microscopic biochemical processes
and local environmental conditions that combine to determine the magnitude
of tbio are independent of the nuclear
astrophysical and
gravitational processes that determine the typical stellar main sequence
lifetime tms. However, this assumption leads to the
striking conclusion
that we should expect extraterrestrial forms of life to be exceptionally
rare [10],
[4],
[11].
The argument, in its simplest form,
is as follows. If tbio and t* are
independent then the time
that life takes to arise is random with respect to the stellar timescale
t*. Thus it is most likely that either
tbio >> t* or that
tbio << t*. Now if tbio
<< t* we must ask why it
is that the first observed inhabited solar system (that is, us) has
tbio 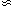 t*. This would be extraordinarily unlikely. On
the other hand, if tbio >> t* then the
first observed inhabited
solar system (us) is most likely to have tbio
t* since
systems with tbio >> t* have yet to
evolve. Thus we are a
rarity, one of the first living systems to arrive on the scene. Generally,
we are led to a conclusion, an extremely pessimistic one for the SETI
enterprise, that tbio >> t*.
t*. This would be extraordinarily unlikely. On
the other hand, if tbio >> t* then the
first observed inhabited
solar system (us) is most likely to have tbio
t* since
systems with tbio >> t* have yet to
evolve. Thus we are a
rarity, one of the first living systems to arrive on the scene. Generally,
we are led to a conclusion, an extremely pessimistic one for the SETI
enterprise, that tbio >> t*.
In order to escape from this conclusion we have to undermine one of the
assumptions underlying the argument that leads to it. For example, if we
suppose that tbio is no independent of
t* then things look
different. If tbio /t* is a rising
function of t* then it is
actually likely that we will find tbio
t*. Livio
[12]
has given a simple model of how it could be that
tbio and t*
are related by a relation of this general form. He takes a very
simple model of the evolution of a life-supporting planetary atmosphere like
the Earth's to have two key phases which lead to its oxygen content:
Phase 1: Oxygen is released by the photodissociation of water vapour. On Earth this took 2.4 x 109 yr and led to an atmospheric O2 build up to about 10-3 of its present value.
Phase 2: Oxygen and ozone levels grow to about 0.1 of their present levels. This is sufficient to shield the Earth's surface from lethal levels of ultra-violet radiation in the 2000-3000 Å band (note that nucleic acid and protein absorption of ultra-violet radiation peaks in the 2600-2700 Å and 2700-2900 Å bands, respectively). On Earth this phase took about 1.6 x 109 yr.
Now the length of Phase 1 might be expected to be inversely proportional to the intensity of radiation in the wavelength interval 1000-2000 Å, where the key molecular levels for H2O absorption lie. Studies of stellar evolution allow us to determine this time interval and provide a rough numerical estimate of the resulting link between the biological evolution time (assuming it to be determined closely by the photodissociation time) and the main sequence stellar lifetime, with [12]
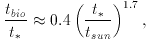
where tsun is the age of the Sun.
This model indicates a possible route to establishing a link between the biochemical timescales for the evolution of life and the astrophysical timescales that determine the time required to create an environment supported by a stable hydrogen burning star. There are obvious weak links in the argument. It provides on a necessary condition for life to evolve, not a sufficient one. We know that there are many other events that need to occur before life can evolve in a planetary system. We could imagine being able to derive an expression for the probability of planet formation around a star. This would involve many other factors which would determine the amount of material available for the formation of solid planets with atmospheres at distances which permit the presence of liquid water and stable surface conditions. Unfortunately, we know that there were many ``accidents'' of the planetary formation process in the solar system which have subsequently played a major role in the existence of long-lived stable conditions on Earth, [13]. For example, the presence of resonances between the precession rates of rotating planets and the gravitational perturbations they feel from all other bodies in their solar system can easily produce chaotic evolution of the tilt of a planet's rotation axis with respect to the orbital plane of the planets over times must shorter than the age of the system [14], [13]. The planet's surface temperature variations, insolation levels, and sea levels are sensitive to this angle of tilt. It determines the climatic differences between what we call ``the seasons''. In the case of the Earth, the modest angle of tilt (approximately 23 degrees) would have experienced this erratic evolution had it not been for the presence of the Moon [15], [13]. The Moon is large enough for its gravitational effects to dominate the resonances which occur between the Earth's precessional rotation and the frequency of external gravitational perturbations from the other planets. As a result the Earth's tilt wobbles only by a fraction of a degree around 23° over hundreds of thousands of years. Enough perhaps to cause some climatic change, but not catastrophic for the evolution of life.
This shows how the causal link between stellar lifetimes and biological evolution times may be rather a minor factor in the chain of fortuitous circumstances that must occur if habitable planets are to form and sustain viable conditions for the evolution of life over long periods of time. The problem remains to determine whether he other decisive astronomical factors in planet formation are functionally linked to the surface conditions needed for biochemical processes.